
One of the important specialized pathways of a number of amino acids is the synthesis of purine and pyrimidine nucleotides. These nucleotides are important for a number of reasons. Most of them, not just ATP, are the sources of energy that drive most of our reactions. ATP is the most commonly used source but GTP is used in protein synthesis as well as a few other reactions. UTP is the source of energy for activating glucose and galactose. CTP is an energy source in lipid metabolism. AMP is part of the structure of some of the coenzymes like NAD and Coenzyme A. And, of course, the nucleotides are part of nucleic acids. Neither the bases nor the nucleotides are required dietary components. (Another perspective on this.) We can both synthesize them de novo and salvage and reuse those we already have.
There are two kinds of nitrogen-containing bases - purines and pyrimidines. Purines consist of a six-membered and a five-membered nitrogen-containing ring, fused together. Pyridmidines have only a six-membered nitrogen-containing ring. There are 4 purines and 4 pyrimidines that are of concern to us.
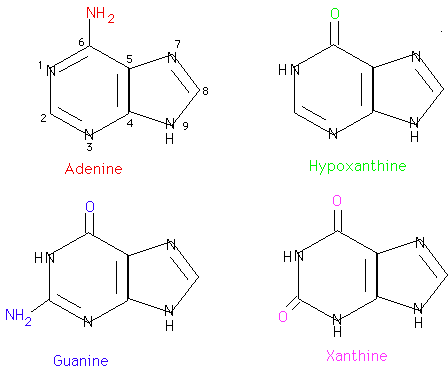
Adenine and guanine are found in both DNA and RNA. Hypoxanthine and xanthine are not incorporated into the nucleic acids as they are being synthesized but are important intermediates in the synthesis and degradation of the purine nucleotides.

Cytosine is found in both DNA and RNA. Uracil is found only in RNA. Thymine is normally found in DNA. Sometimes tRNA will contain some thymine as well as uracil.
If a sugar, either ribose or 2-deoxyribose, is added to a nitrogen base, the resulting compound is called a nucleoside. Carbon 1 of the sugar is attached to nitrogen 9 of a purine base or to nitrogen 1 of a pyrimidine base. The names of purine nucleosides end in -osine and the names of pyrimidine nucleosides end in -idine. The convention is to number the ring atoms of the base normally and to use l', etc. to distinguish the ring atoms of the sugar. Unless otherwise specificed, the sugar is assumed to be ribose. To indicate that the sugar is 2'-deoxyribose, a d- is placed before the name.

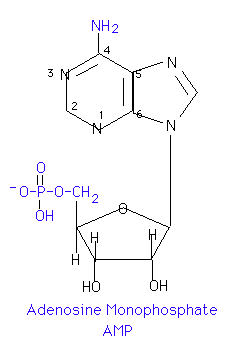
Adding one or more phosphates to the sugar portion of a nucleoside results in a nucleotide. Generally, the phosphate is in ester linkage to carbon 5' of the sugar. If more than one phosphate is present, they are generally in acid anhydride linkages to each other. If such is the case, no position designation in the name is required. If the phosphate is in any other position, however, the position must be designated. For example, 3'-5' cAMP indicates that a phosphate is in ester linkage to both the 3' and 5' hydroxyl groups of an adenosine molecule and forms a cyclic structure. 2'-GMP would indicate that a phosphate is in ester linkage to the 2' hydroxyl group of a guanosine. Some representative names are:
Nucleotides are joined together by 3'-5' phosphodiester bonds to form
polynucleotides. Polymerization of ribonucleotides will produce an RNA while
polymerization of deoxyribonucleotides leads to DNA. 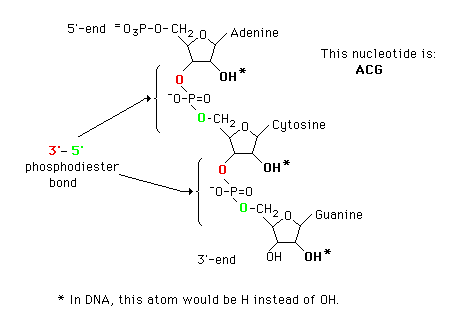
Most, but not all, nucleic acids in the cell are associated with protein. Dietary nucleoprotein is degraded by pancreatic enzymes and tissue nucleoprotein by lysosomal enzymes. After dissociation of the protein and nucleic acid, the protein is metabolized like any other protein.
The nucleic acids are hydrolyzed randomly by nucleases to yield a mixture of polynucleotides. These are further cleaved by phosphodiesterases (exonucleases) to a mixture of the mononucleotides. The specificity of the pancreatic nucleotidases gives the 3'-nucleotides and that of the lysosomal nucleotidases gives the biologically important 5'-nucleotides.
The nucleotides are hydrolyzed by nucleotidases to give the nucleosides and Pi. This is probably the end product in the intestine with the nucleosides being the primary form absorbed. In at least some tissues, the nucleosides undergo phosphorolysis with nucleoside phosphorylases to yield the base and ribose 1-P (or deoxyribose 1-P). Since R 1-P and R 5-P are in equilibrium, the sugar phosphate can either be reincorporated into nucleotides or metabolized via the Hexose Monophosphate Pathway. The purine and pyrimidine bases released are either degraded or salvaged for reincorporation into nucleotides. There is significant turnover of all kinds of RNA as well as the nucleotide pool. DNA doesn't turnover but portions of the molecule are excised as part of a repair process.

Purine and pyrimidines from tissue turnover which are not salvaged are catabolized and excreted. Little dietary purine is used and that which is absorbed is largely catabolized as well. Catabolism of purines and pyrimidines occurs in a less useful fashion than did the catabolism of amino acids in that we do not derive any significant amount of energy from the catabolism of purines and pyrimidines. Pyrimidine catabolism, however, does produce beta-alanine, and the endproduct of purine catabolism, which is uric acid in man, may serve as a scavenger of reactive oxygen species.
Purine Catabolism
The end product of purine catabolism in man is uric acid. Other
mammals have the
enzyme urate oxidase and excrete the more soluble allantoin as the end product.
Man does not have this enzyme so urate is the end product for us. Uric acid is
formed primarily in the liver and excreted by the kidney into the urine.

Guanine nucleotides are hydrolyzed to the nucleoside guanosine which undergoes phosphorolysis to guanine and ribose 1-P. Man's intracellular nucleotidases are not very active toward AMP, however. Rather, AMP is deaminated by the enzyme adenylate (AMP) deaminase to IMP. In the catobilsm of purine nucleotides, IMP is further degraded by hydrolysis with nucleotidase to inosine and then phosphorolysis to hypoxanthine.
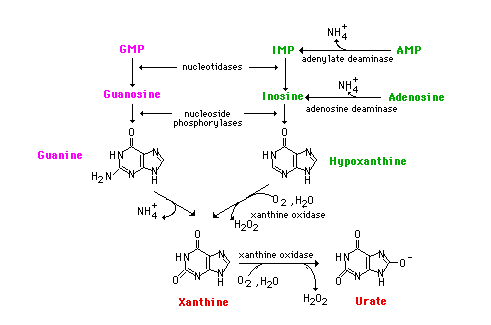
Adenosine does occur but usually arises from S-Adenosylmethionine during the course of transmethylation reactions. Adenosine is deaminated to inosine by an adenosine deaminase. Deficiencies in either adenosine deaminase or in the purine nucleoside phosphorylase lead to two different immunodeficiency diseases by mechanisms that are not clearly understood. With adenosine deaminase deficiency, both T and B-cell immunity is affected. The phosphorylase deficiency affects the T cells but B cells are normal. In September, 1990, a 4 year old girl was treated for adenosine deaminase deficiency by genetically engineering her cells to incorporate the gene. The treatment,so far, seems to be successful.
Whether or not methylated purines are catabolized depends upon the location of the methyl group. If the methyl is on an -NH2, it is removed along with the -NH2 and the core is metabolized in the usual fashion. If the methyl is on a ring nitrogen, the compound is excreted unchanged in the urine.
Both adenine and guanine nucleotides converge at the common intermediate xanthine. Hypoxanthine, representing the original adenine, is oxidized to xanthine by the enzyme xanthine oxidase. Guanine is deaminated, with the amino group released as ammonia, to xanthine. If this process is occurring in tissues other than liver, most of the ammonia will be transported to the liver as glutamine for ultimate excretion as urea.
Xanthine, like hypoxanthine, is oxidized by oxygen and xanthine oxidase with the production of hydrogen peroxide. In man, the urate is excreted and the hydrogen peroxide is degraded by catalase. Xanthine oxidase is present in significant concentration only in liver and intestine. The pathway to the nucleosides, possibly to the free bases, is present in many tissues.
Both undissociated uric acid and the monosodium salt (primary form in blood) are only sparingly soluble. The limited solubility is not ordinarily a problem in urine unless the urine is very acid or has high [Ca2+]. [Urate salts coprecipitate with calcium salts and can form stones in kidney or bladder.] A very high concentration of urate in the blood leads to a fairly common group of diseases referred to as gout. The incidence of gout in this country is about 3/1000.
Gout is a group of pathological conditions associated with markedly elevated levels of urate in the blood (3-7 mg/dl normal). Hyperuricemia is not always symptomatic, but, in certain individuals, something triggers the deposition of sodium urate crystals in joints and tissues. In addition to the extreme pain accompanying acute attacks, repeated attacks lead to destruction of tissues and severe arthritic-like malformations. The term gout should be restricted to hyperuricemia with the presence of these tophaceous deposits.
Urate in the blood could accumulate either through an overproduction and/or an underexcretion of uric acid. In gouts caused by an overproduction of uric acid, the defects are in the control mechanisms governing the production of - not uric acid itself - but of the nucleotide precursors. The only major control of urate production that we know so far is the availability of substrates (nucleotides, nucleosides or free bases).
One approach to the treatment of gout is the drug allopurinol, an
isomer of
hypoxanthine. 
Allopurinol is a substrate for xanthine oxidase, but the product binds so tightly that the enzyme is now unable to oxidized its normal substrate. Uric acid production is diminished and xanthine and hypoxanthine levels in the blood rise. These are more soluble than urate and are less likely to deposit as crystals in the joints. Another approach is to stimulate the secretion of urate in the urine.
In summary, all, except ring-methylated, purines are deaminated (with the amino group contributing to the general ammonia pool) and the rings oxidized to uric acid for excretion. Since the purine ring is excreted intact, no energy benefit accrues to man from these carbons.
Pyrimidine Catabolism
In contrast to purines, pyrimidines undergo ring cleavage and the usual end
products of catabolism are beta-amino acids plus ammonia and carbon dioxide.
Pyrimidines from nucleic acids or the energy pool are acted upon by
nucleotidases
and pyrimidine nucleoside phosphorylase to yield the free bases. The
4-amino group
of both cytosine and 5-methyl cytosine is released as ammonia.
In order for the rings to be cleaved, they must first be reduced by NADPH. Atoms 2 and 3 of both rings are released as ammonia and carbon dioxide. The rest of the ring is left as a beta-amino acid. Beta-amino isobutyrate from thymine or 5-methyl cytosine is largely excreted. Beta-alanine from cytosine or uracil may either be excreted or incorporated into the brain and muscle dipeptides, carnosine (his-beta-ala) or anserine (methyl his-beta-ala).

Purine and pyrimidine bases which are not degraded are recycled - i.e. reincorporated into nucleotides. This recycling, however, is not sufficient to meet total body requirements and so some de novo synthesis is essential. There are definite tissue differences in the ability to carry out de novo synthesis. De novo synthesis of purines is most active in liver. Non-hepatic tissues generally have limited or even no de novo synthesis. Pyrimidine synthesis occurs in a variety of tissues. For purines, especially, non-hepatic tissues rely heavily on preformed bases - those salvaged from their own intracellular turnover supplemented by bases synthesized in the liver and delivered to tissues via the blood.
"Salvage" of purines is reasonable in most cells because xanthine oxidase, the key enzyme in taking the purines all of the way to uric acid, is significantly active only in liver and intestine. The bases generated by turnover in non-hepatic tissues are not readily degraded to uric acid in those tissues and, therefore, are available for salvage. The liver probably does less salvage but is very active in de novo synthesis - not so much for itself but to help supply the peripheral tissues.
De novo synthesis of both purine and pyrimidine nucleotides occurs from readily available components.
De Novo Synthesis of Purine Nucleotides
We use for purine nucleotides the entire glycine molecule (atoms 4, 5,7), the amino nitrogen of aspartate (atom 1), amide nitrogen of glutamine (atoms 3, 9), components of the folate-one-carbon pool(atoms 2, 8), carbon dioxide, ribose 5-P from glucose and a great deal of energy in the form of ATP. In de novo synthesis, IMP is the first nucleotide formed. It is then converted to either AMP or GMP.
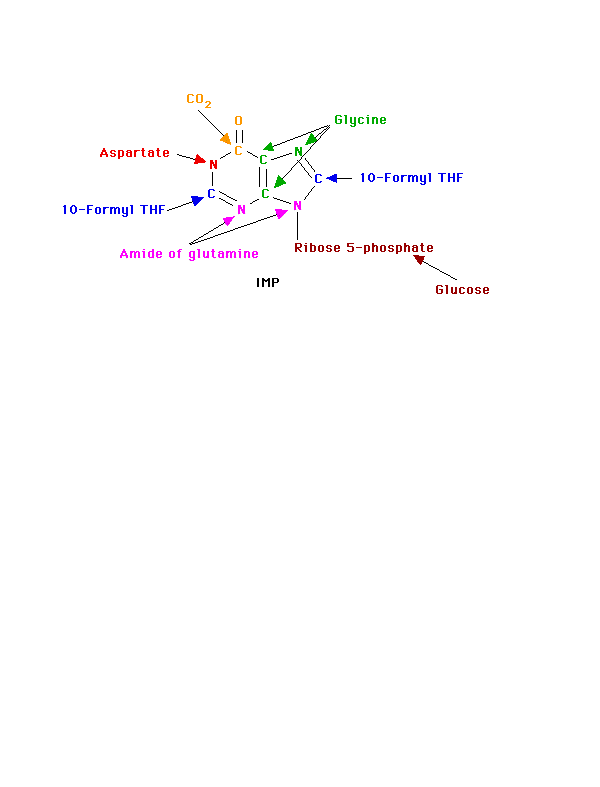
Since the purines are synthesized as the ribonucleotides, (not as the free bases) a necessary prerequisite is the synthesis of the activated form of ribose 5-phosphate. Ribose 5-phosphate reacts with ATP to form 5-Phosphoribosyl-1-pyrophosphate (PRPP).

This reaction occurs in many tissues because PRPP has a number of roles - purine and pyrimidine nucleotide synthesis, salvage pathways, NAD and NADP formation. The enzyme is heavily controlled by a variety of compounds (di- and tri-phosphates, 2,3-DPG), presumably to try to match the synthesis of PRPP to a need for the products in which it ultimately appears.
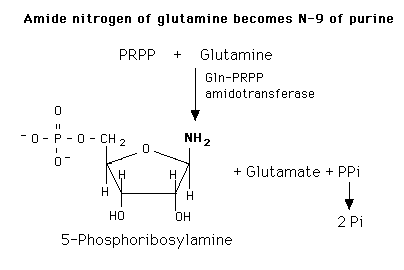
De novo purine nucleotide synthesis occurs actively in the cytosol of the liver where all of the necessary enzymes are present as a macro-molecular aggregate. The first step is a replacement of the pyrophosphate of PRPP by the amide group of glutamine. The product of this reaction is 5-Phosphoribosylamine. The amine group that has been placed on carbon 1 of the sugar becomes nitrogen 9 of the ultimate purine ring. This is the commitment and rate-limiting step of the pathway.
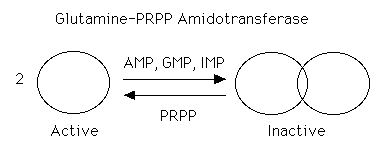
The enzyme is under tight allosteric control by feedback inhibition. Either AMP, GMP, or IMP alone will inhibit the amidotransferase while AMP + GMP or AMP + IMP together act synergistically. This is a fine control and probably the major factor in minute by minute regulation of the enzyme. The nucleotides inhibit the enzyme by causing the small active molecules to aggregate to larger inactive molecules.
[PRPP] also can play a role in regulating the rate. Normal intracellular concentrations of PRPP (which can and do fluctuate) are below the KM of the enzyme for PRPP so there is great potential for increasing the rate of the reaction by increasing the substrate concentration. The kinetics are sigmoidal. The enzyme is not particularly sensitive to changes in [Gln] (Kinetics are hyperbolic and [gln] approximates KM). Very high [PRPP] also overcomes the normal nucleotide feedback inhibition by causing the large, inactive aggregates to dissociate back to the small active molecules.
Purine de novo synthesis is a complex, energy-expensive pathway. It should be, and is, carefully controlled.
Once the commitment step has produced the 5-phosphoribosyl amine, the rest of the molecule is formed by a series of additions to make first the 5- and then the 6-membered ring. (Note: the numbers given to the atoms are those of the completed purine ring and names, etc. of the intermediate compounds are not given.) The whole glycine molecule, at the expense of ATP adds to the amino group to provide what will eventually be atoms 4, 5, and 7 of the purine ring (The amino group of 5-phosphoribosyl amine becomes nitrogen N of the purine ring.) One more atom is needed to complete the five-membered ring portion and that is supplied as 5, 10-Methenyl tetrahydrofolate.
Before ring closure occurs, however, the amide of glutamine adds to carbon 4 to start the six-membered ring portion (becomes nitrogen 3). This addition requires ATP. Another ATP is required to join carbon 8 and nitrogen 9 to form the five-membered ring.
The next step is the addition of carbon dioxide (as a carboxyl group) to form carbon 6 of the ring. The amine group of aspartate adds to the carboxyl group with a subsequent removal of fumarate. The amino group is now nitrogen 1 of the final ring. This process, which is typical for the use of the amino group of aspartate, requires ATP. The final atom of the purine ring, carbon 2, is supplied by 10-Formyl tetrahydrofolate. Ring closure produces the purine nucleotide, IMP.
Note that at least 4 ATPs are required in this part of the process. At no time do we have either a free base or a nucleotide.

IMP can then become either AMP or GMP. GMP formation requires that IMP be first oxidized to XMP using NAD. The oxygen at position 2 is substituted by the amide N of glutamine at the expense of ATP. Similarly, GTP provides the energy to convert IMP to AMP. The amino group is provided by aspartate in a mechanism similar to that used in forming nitrogen 1 of the ring. Removal of the carbons of aspartate as fumarate leaves the nitrigen behind as the 6-amino group of the adenine ring. The monophosphates are readily converted to the di- and tri-phosphates.

Control of purine nucleotide synthesis has two phases. Control of the synthesis as a whole occurs at the amidotransferase step by nucleotide inhibition and/or [PRPP]. The second phase of control is involved with maintaining an appropriate balance (not equality) between ATP and GTP. Each one stimulates the synthesis of the other by providing the energy. Feedback inhibition also controls the branched portion as GMP inhibits the conversion of IMP to XMP and AMP inhibits the conversion of IMP to adenylosuccinate.

Possible Scenario:
One could imagine the controls operating in such a way that if only one of the two nucleotides were required, there would be a partial inhibition of de novo synthesis because of high levels of the other and the IMP synthesized would be directed toward the synthesis of the required nucleotide. If both nucleotides were present in adequate amounts, their synergistic effect on the amidotransferase would result in almost complete inhibition of de novo synthesis.
De Novo Synthesis of Pyrimidine Nucleotides
Since pyrimidine molecules are simpler than purines, so is their synthesis simpler but is still from readily available components. Glutamine's amide nitrogen and carbon dioxide provide atoms 2 and 3 or the pyrimidine ring. They do so, however, after first being converted to carbamoyl phosphate. The other four atoms of the ring are supplied by aspartate. As is true with purine nucleotides, the sugar phosphate portion of the molecule is supplied by PRPP.

Pyrimidine synthesis begins with carbamoyl phosphate synthesized in the cytosol of those tissues capable of making pyrimidines (highest in spleen, thymus, GItract and testes). This uses a different enzyme than the one involved in urea synthesis. Carbamoyl phosphate synthetase II (CPS II) prefers glutamine to free ammonia and has no requirement for N-Acetylglutamate.

Carbamoyl phosphate condenses with aspartate in the presence of aspartate transcarbamylase to yield N-carbamylaspartate which is then converted to dihydroorotate.
In man, CPSII, asp-transcarbamylase, and dihydroorotase activities are part of a multifunctional protein.
Oxidation of the ring by a complex, poorly understood enzyme produces the free pyrimidine, orotic acid. This enzyme is located on the outer face of the inner mitochondrial membrane, in contrast to the other enzymes which are cytosolic. Note the contrast with purine synthesis in which a nucleotide is formed first while pyrimidines are first synthesized as the free base.

Orotic acid is converted to its nucleotide with PRPP. OMP is then converted sequentially - not in a branched pathway - to the other pyrimidine nucleotides. Decarboxylation of OMP gives UMP. O-PRT and OMP decarboxylase are also a multifunctional protein. After conversion of UMP to the triphosphate, the amide of glutamine is added, at the expense of ATP, to yield CTP.

The control of pyrimidine nucleotide synthesis in man is exerted primarily at the level of cytoplasmic CPS II. UTP inhibits the enzyme, competitively with ATP. PRPP activates it. Other secondary sites of control also exist (e.g. OMP decarboxylase is inhibited by UMP and CMP). These are probably not very important under normal circumstances.
In bacteria, aspartate transcarbamylase is the control enzyme. There is only one carbamoyl phosphate synthetase in bacteria since they do not have mitochondria. Carbamoyl phosphate, thus, participates in a branched pathway in these organisms that leads to either pyrimidine nucleotides or arginine.
Interconversion of Nucleotides
The monophosphates are the forms synthesized de novo although the triphosphates are the most commonly used forms. But, of course, the three forms are in equilibrium. There are several enzymes classified as nucleoside monophosphate kinases which catalyze the general reaction:(= represents a reversible reaction)
Base-monophosphate + ATP = Base-diphosphate + ADP
e.g. Adenylate kinase: AMP + ATP = 2 ADP
There is a different enzyme for GMP, one for pyrimidines and also enzymes that recognize the deoxy forms.
Similarly, the diphosphates are converted to the triphosphates by nucleoside diphosphate kinase:
BDP + ATP = BTP + ADP
There may be only one nucleoside diphosphate kinase with broad specificity. One can legitimately speak of a pool of nucleotides in equilibrium with each other.
Salvaging of purine and pyrimidine bases is an exceedingly important process for most tissues. There are two distinct pathways possible for salvaging the bases.
The more important of the pathways for salvaging purines uses enzymes called phosphoribosyltransferases (PRT):
PRTs catalyze the addition of ribose 5-phosphate to the base from PRPP to yield a nucleotide.:
Base + PRPP = Base-ribose-phosphate (BMP) + PPi
We gave already seen one example of this type of enzyme as a normal part of de novo synthesis of the pyrimidine nucleotides, - O-PRT.
As a salvage process though, we are dealing with purines. There are two enzymes, A-PRT and HG-PRT. A-PRT is not very important because we generate very little adenine. (Remember that the catabolism of adenine nucleotides and nucleosides is through inosine). HG-PRT, though, is exceptionally important and it is inhibited by both IMP and GMP. This enzyme salvages guanine directly and adenine indirectly. Remember that AMP is generated primarily from IMP, not from free adenine.
HG-PRT is deficient in the disease called Lesch-Nyhan Syndrome, a severe neurological disorder whose most blatant clinical manifestation is an uncontrollable self-mutilation. Lesch-Nyhan patients have very high blood uric acid levels because of an essentially uncontrolled de novo synthesis. (It can be as much as 20 times the normal rate). There is a significant increase in PRPP levels in various cells and an inability to maintain levels of IMP and GMP via salvage pathways. Both of these factors could lead to an increase in the activity of the amidotransferase.
A second type of salvage pathway involves two steps and is the major pathway for the pyrimidines, uracil and thymine.
Base + Ribose 1-phosphate = Nucleoside + Pi (nucleoside phosphorylase)
Nucleoside + ATP - Nucleotide + ADP (nucleoside kinase - irreversible)
There is a uridine phosphorylase and kinase and a deoxythymidine phosphorylase and a thymidine kinase which can salvage some thymine in the presence of dR 1-P.
Formation of Deoxyribonucleotides
De novo synthesis and most of the salvage pathways involve the ribonucleotides. (Exception is the small amount of salvage of thymine indicated above.) Deoxyribonucleotides for DNA synthesis are formed from the ribonucleotide diphosphates (in mammals and E. coli).
A base diphosphate (BDP) is reduced at the 2' position of the ribose portion using the protein, thioredoxin and the enzyme nucleoside diphosphate reductase. Thioredoxin has two sulfhydryl groups which are oxidized to a disulfide bond during the process. In order to restore the thioredoxin to its reduced for so that it can be reused, thioredoxin reductase and NADPH are required.
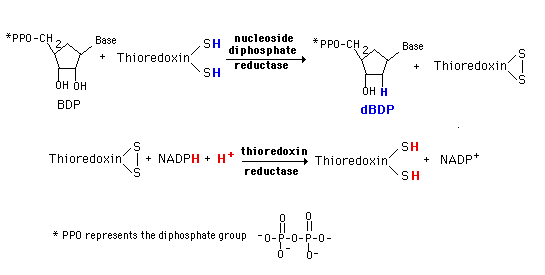
This system is very tightly controlled by a variety of allosteric effectors. dATP is a general inhibitor for all substrates and ATP an activator. Each substrate then has a specific positive effector (a BTP or dBTP). The result is a maintenance of an appropriate balance of the deoxynucleotides for DNA synthesis.
DNA synthesis also requires dTMP (dTTP). This is not synthesized in the de novo pathway and salvage is not adequate to maintain the necessary amount. dTMP is generated from dUMP using the folate-dependent one-carbon pool.
Since the nucleoside diphosphate reductase is not very active toward UDP, CDP is reduced to dCDP which is converted to dCMP. This is then deaminated to form dUMP. In the presence of 5,10-Methylene tetrahydrofolate and the enzyme thymidylate synthetase, the carbon group is both transferred to the pyrimidine ring and further reduced to a methyl group. The other product is dihydrofolate which is subsequently reduced to the tetrahydrofolate by dihydrofolate reductase.

Thymidylate synthetase is particularly sensitive to availability of the folate one-carbon pool. Some of the cancer chemotherapeutic agents interfere with this process as well as with the steps in purine nucleotide synthesis involving the pool.
Cancer chemotherapeutic agents like methotrexate (4-amino, 10-methyl folic acid) and aminopterin (4-amino, folic acid) are structural analogs of folic acid and inhibit dihydrofolate reductase. This interferes with maintenance of the folate pool and thus of de novo synthesis of purine nucleotides and of dTMP synthesis. Such agents are highly toxic and administered under careful control.
If you would like to test your level of understanding, you may try these multiple choice quiz questions.
Return to the NetBiochem Welcome page, where you can choose another topic.
Last modified 12/4/97
